| | | |
 Home /
Home /
Death Squared 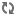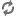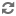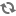 Uploading .... Uploading ....
Introduction to Death Squared
by
Ed Coll 5-15-96
Living on an island like Kauai my thoughts often wander to questions
of sustainability, quality of life, and carrying capacity. When Captain
Cook came to Kauai there were an estimated 50,000 Native Hawaiians living
on Kauai. That's about the population of Kauai today. But there is a difference.
Today, Kauai's 50,000 residents are 85% dependent on imports. My definition
of "carrying capacity" for Kauai is the number of people this
island can support on a sustainable basis without the importation of goods.
"We can't alter our lifestyle and go back to living in grass shacks.
We've got to be good consumers to keep the GNP rolling", an anonymous
source says.
Well, what's the alternative?
What if Kauai was a true paradise with all our housing, food, and water
provided for us in abundance? What if more and more people moved to Kauai?
At what point would the population density turn paradise into a living hell?
Death Squared is the story of a paradise for mice called Universe 25. All
the needs of the mice were provided for (shelter, food, water, etc.). Four
pairs of mice colonized Universe 25 and began breeding. The population increased
with some interesting results regarding survival. This is not a science
fiction story but a real scientific study completed in the 1970's. Is there a lesson for humankind
in this tale of mice?
Read on but please keep in mind this mantra: "People aren't mice. We
can adapt."
Can we?
Death Squared:
The Explosive Growth and Demise of a Mouse Population
by John B Calhoun MD
(Section on Behavioral Systems,
Laboratory of Brain Evolution & Behavior,
National Institute of Mental Health,
9000 Rockville Pike,
Bethesda, Maryland 20014, USA)
Fig 1 Universe 25,
February 10,1970, 681 days after colonization. Calhoun in universe with
an exceptionally large aggregate of pooled withdrawn mice on the floor just
across the centre of the floor from him. Note the aggregation of mice on
some food hoppers in contrast to their near absence on others. This is the
'behavioural sink' phenomenon in which the learned need for proximity
to others as a secondary reinforcer at a resource site gains dominance over
the primary need, in this case food.
- Photo by Yoichi R Okamoto -
I shall largely speak of mice, but my thoughts are on man, on healing,
on life and its evolution. Threatening life and evolution are the two death:
death of the spirit and death of the body. Evolution, in terms of ancient
wisdom, is the acquisition of access to the tree of life. This takes us
back to the white first horse of the Apocalypse which with its rider set
out to conquer the forces that threaten the spirit with death. Fourth: in
Revelations (liabilities) we note: 'To him who conquers I will grant to
eat the tree of life, which is in the paradise of God' and further on (Rev.
xxii.2): 'The leaves of the tree were for the healing of nations.'
This takes us to the fourth horse of the Apocalypse (Rev. vi.7): 'I saw...
a pale horse, and its rider's name was Death, and Hades followed him; and
they were given power over a fourth of the earth, to kill with the sword
and with famine and with pestilence and by wild beasts of the earth' (italics
mine). This second death has gradually become the predominant concern of
modern medicine. And yet there is nothing in the earlier history of medicine,
or in the precepts embodied in the Hippocratic Oath, that precludes medicine
from being equally concerned with healing the spirit, and healing nations,
as with healing the body. Perhaps we might do well to reflect upon another
of John's transcriptions (Rev. ii.ll): 'He who conquers shall not be hurt
by the second death.'
Bodily Mortality
Let us first consider the second death Table
I). The four mortality factors listed in
Revelation have direct counterparts (with a division of one of them to form
a total of five) in the ecology of animals in nature. I shall briefly treat
each of these five mortality factors, and then discuss the steps taken to
eliminate, or drastically reduce, the impact of each in a Utopian
environment constructed for mice.
(1) Emigration: Animals in the wild rarely die from the 'sword' directly;
few are the deaths from intraspecific combat Rather it is that individuals,
who have failed in the more symbolic conflicts involved in gaining the right
to remain in the locality of their birth or in more preferred habitats,
take recourse to movement into peripheral unoccupied or suboptimal habitats
In strange and less favourable habitats the emigres become more exposed
to other mortality factors. Any removal of excess members from an established
population, from the point of view of change in its numbers, represents
mortality.In this sense emigration is a mortality factor.
(2a) Resource shortage: classically food and water have received
the focus of attention with respect to their reduction increasing the likelihood
of death. Shortages of shelter, other environmental resources and associates
lead to debilitation, and an unsatisfactoriness of habitat that culminate
in death or failure to reproduce (species death).
(2b.) Inclement weather: Every species of animal has developed a
genetic adaptation to some particular range of external conditions which
affect its physiology. Any conditions of wind, rain, humidity or temperature
which exceed the usual limits of tolerance bring death immediately or increase
the risk of death through debilitation. Beyond these more usual contributions
to mortality, flood and fire represent cataclysmic changes that have widespread
and more long-lasting effects on population numbers.
(3) Disease: Although most animals develop some capacity to tolerate
the parasites, bacteria and viruses that invade their bodies, most species
still remain subject to epidemic decimation in addition to a more normal
attrition from disease. Abnormally high densities enhance the likelihood
of spread of disease to epidemic proportions.
(4) Predation: Practically every species has, through evolution,
had associated with it predators capable of killing some of its members.
Despite loss of members from these five kinds of mortality, most species
persist over long Periods, even those measured in geological time. To do
so every such species has developed capacities for reproduction requisite
to compensate for losses from the normal impact of mortality factors other
than aging. I have omitted senescence as a mortality factor since rare
is the animal who lives long enough in nature to reach a post-reproductive
age without having succumbed to one of the above mortality factors.
A Mortality-inhibiting Environment for Mice
Some of the attributes of this environment have been discussed or figured
elsewhere (Calhoun 1969, 1971, Wigotsky 1970). Here I shall describe how
its design reduced mortality.
(1) Emigration prevention: A closed physical universe was formed
by four 54 inch (1.37 m) high walls forming a square of side 101 inches
(2.57 m). Although the walls were structured for use by mice to increase
the effective use-area of the universe, the mice could not climb over the
upper 17 inch (43 cm) unstructured portion of the galvanized metal walls.
(2a) Resource supra-availability: Each 25* inch (64 cm) linear segment
of wall was identically structured. Four 3 inch (7.6 cm) diameter tunnels
34 inches (86 cm) long of j inch (12.5 mm) mesh wire were soldered vertically
to the walls. The open lower end, just above the floor which was covered
with ground corn cob, gave access to each tunnel. At 8 inch (20.3 cm) intervals
above the floor in each tunnel four 1 * inch (3.2 cm) openings through the
tunnel mesh and the metal wall gave access to 8x5x4 inch (20.3x12.7x 10.2
cm) retreat nesting boxes. Fifteen mice could comfortably rest in a single
nest box.
Thus there were four
four-unit walk-up
one-room apartments
in each cell.
Thus there were four four-unit walk-up one-room apartments in each cell.
The cell is the replicated wall configuration, here being described, with
an associated 640 sq. inch (0.356 m2) of floor space. A wire mesh food hopper
with a 6 x 10 inch (15x25 cm) surface was located in each of the 16 cells
halfway up the wall and in contact with the right-hand tunnel of each set
of four tunnels. Mice climbing across the outer surface of the tunnel gained
access to the food hopper. Twenty five mice could feed simultaneously on
a hopper. By further climbing up the outside surface of the tunnels mice
had access to a 4x18 inch (10x45 cm) platform above which four water bottles
were suspended. Two mice could drink simultaneously at each bottle. An abundant
supply of paper strips for nesting material was always available on the
floor a few inches out from the bases of the tunnels. Considering the time
required to eat and drink, access to food would not have been a limiting
variable until a population of 9500 was reached, or 6144 for water. Considering
that there were 256 nest retreat sites in the 16 cells one would not expect
shelter to be a limiting factor until the population exceeded 3840. Due
to the tendency of many animals to choose to crowd together in numbers in
excess of 15 per nest site, at the peak population size of 2200 mice, 20%
of all nest sites were usually unoccupied. Thus there was always opportunity
for females to select an unoccupied space for rearing young if they so chose.
(2b) Weather amelioration: The mouse universe was located on the
second floor of a prefabricated metal building. During the cooler months
of the year ambient temperature was kept close to 68F (20C) During the warmer
months of the year large exhaust fans kept ambient temperatures mostly within
the 70-90F (21-32C) range of the outside environment. Being indoors, rain
could not contribute to debilitation. Air movement was kept low, except
when this favoured heat loss during periods of higher ambient temperature.
No evidence was ever obtained to indicate that such weather conditions enhanced
mortality.
(3) Disease control: The Balb C albino house mice (Mus musculus)
used as colonizers in this study were obtained from the National Institutes
of Health breeding colony where extreme precautions are followed to preclude
establishment of epidemic type diseases such as salmonella. Bacterial culture
taken at the highest density of the population indicated that such organisms
were not a factor in our study. About every four to eight weeks the ground
corn cob in all nest boxes and the floor. was removed along with accumulated
feces.
(4) Predation: No predators were present.
Some mortality did occur throughout the history of the population we initiated
in this 16 cell universe. Not until the mice became quite old did the mortality
from ageing contribute significantly to removal of members from the population.
On the basis of initial analyses, menopause in females comes at about 560
days of age. Although we have not yet determined the average life expectancy
at weaning, I suspect that it is well past menopause. Large numbers of mice
lived to 800 days of age, which is equivalent to 80 years of age for a human.
Explosive Initial Population Growth, the Resource Exploitation Phase
B
Four pairs of 48-day-old Balb C strain house mice were introduced into the
16 cell universe on July 9 1968, after each mouse had been isolated for
21 days following weaning. There followed a period of 104 days (Phase A)
before the first litters were born. These 104 days were marked by considerable
social turmoil among these 8 mice until they became adjusted to each other
and to their expanded surroundings. Following this adjustment and the birth
of the first litters the population exhibited an exponential increase, with
a doubling time of about 55 days (Fig 2). This progression of numbers was approximately 20,
40, 80, 160, 320, to 620 after nearly five doublings. I call this period
of most rapid growth Phase B. At 620 weaned mice the rate of population
growth abruptly decreased to a doubling time of approximately 145 days.
Periodically through Phase B, young born into the universe reached sexual
maturity and bore young, thus contributing to the compound interest rate
of population growth.
Distribution of place of birth (Fig 3) of mice born during Phase B provides an insight into
the social organization that developed. The square represents the wall of
the pen, and the small black rectangles depict the locations of the food
hoppers along its inner surface. Between each two hoppers there are four
sets of walk-up apartments, each set containing four nest boxes accessible
by a single tunnel. The total young born in each of the 64 sets of four
nest boxes were tabulated for the period through the first survey after
the termination of Phase B. These totals are shown in Fig
3 as open bars extending outward from the wall of the pen.
It may be seen that births tended to be concentrated in some sets of nest
boxes, while others had few or none. This uneven distribution of births
reflects a clustering of reproducing females into brood groups defined in
Fig 3 by lines radiating toward the centre of
the universe from the bases of nest boxes at the interface between each
two adjoining brood groups. Total births per brood group are indicated between
radial lines. These totals reflect two properties of a closed social system:
(1) Bilateral symmetry: The northeast brood group produced only 13
young in 252 days whereas the opposite southwest brood group produced over
eight times as many, 111. Between these two extremes from the location of
the peak producing brood group toward the least productive one, in both
clockwise and counterclockwise directions, there is a decline in number
of young born. This trend reflects the attempt by the members of the population
to superimpose a more effective bilateral symmetry of organization upon
an environment that tends toward radial symmetry.
(2) Hierarchy of groups: Productivity of a group may be taken as
an index of its social status. The fourteen brood groups may thus be ranked
with rank No.1 assigned to the group which produced 111 young, and rank
No.14 to the one which produced only 13 young. Plotting the number of young
produced as a function of this ranking (Fig 4) reflects a remarkable
hierarchical ordering within the total social system. This type of ordering
is identical to that which results when we examine the degree of activity
exhibited by the several males of an interacting group. The most dominant
male is the most active one, and as social dominance declines so does the
degree of activity. Such activity is termed 'social velocity' (Calhoun 1963,
1967, 1971). Our studies show that in a group of 14 males social velocity
declines linearly with rank with approximately the same slope as in Fig
4. Each of the brood groups had associated with it a male which was
territorially dominant within an area on the floor roughly coinciding with
the sectors shown in Fig 3. All the ranges of these territorial males overlapped
near the centre of the universe The most dominant male was always associated
with the brood group that produced the most young, and the degree of dominance
of the other territorial males tended to be reflected by the productivity
of females associated with them.
Both bilateral and hierarchical social organization during Phase B contributed
to a maximum exploitation of resources that led to an explosive rate of
increase of the population. At the end of this phase all the most desirable
physical space was filled with organized social groups. These 14 social
groups totaled 150 adults. On average each group contained over 10 individuals
including a territorial male, associated males and females and their juvenile
and subadult progeny. At the end of Phase B there were 470 of these immature
mice that had experienced good maternal care and early socialization. Thus
there were over three times as many younger animals as the socially established
older ones. This number is far greater than would have existed had the normal
ecological mortality factors functioned.
Inhibited Secondary Population Growth, the Stagnation Phase C
Beginning at Day 315 after colonization and continuing for 245 more days,
the population grew at a much slower rate, doubling only every 145 days
rather than each 55 days as in Phase B. let us examine the circumstances
surrounding this decline in rate of population growth. In the normal course
of events in a natural ecological setting somewhat more young survive to
maturity than are necessary to replace their dying or senescent established
associates. The excess that find no social niches emigrate. However, in
my experimental universe there was no opportunity for emigration. As the
unusually large number of young gained adulthood they had to remain, and
they did contest for roles in the filled social system. Males who failed
withdrew physically and psychologically; they became very inactive and aggregated
in large pools near the centre of the floor of the universe. From this point
on they no longer initiated interaction with their established associates,
nor did their behavior elicit attack by territorial males. Even so, they
became characterized by many wounds and much scar tissue as a result of
attacks by other withdrawn males. Return of 2 or more males, who had gone
to eat and drink, marked an abrupt shift in the level of ambient stimuli
for their quiescent associates. Resultant excitation often precipitated
one of the resting males into an attack upon his other withdrawn associates
who, having lost the capacity for fleeing remained relatively immobile despite
receiving vicious attacks. A mouse so attacked would at a later time become
an attacker. Female counterparts of these withdrawn males tended to withdraw
to higher level boxes that were less preferred by females with litters.
Such females were not characterized by the violent aggression of the withdrawn
males.
As a result of the extreme demands made on territorial males to reject maturing
associates, their ability to continue territorial defence decline. Gradually
the frequency of this involvement in territorial defence declined as did
the area defended. This left nursing females more exposed to invasion of
their nest sites. Normally nursing females in the presence of territorial
males exhibit little aggression. However, in response to invasion of nest
sites and bases of ramps leading to them, the nursing females did become
aggressive, essentially taking over the role of the territorial males. This
aggression generalized to their own young who were attacked, wounded, and
forced to leave home several days before normal weaning. During Phase C
the incidence of conception declined, and resorption of fetuses increased.
Maternal behaviour also became disrupted. Young were often wounded in the
delivery process. Females transported their young to several sites, during
which process some were abandoned. Many litters of a young age on one survey
disappeared before the next survey. Such abandoning of young following survey
disturbance is a particularly sensitive index of dissolution of maternal
behaviour. The combined effect of these several factors affecting reduced
conception, increased fetal mortality and increased preweaning mortality
largely accounts for the abrupt decline in rate of population growth characterizing
Phase C. For all practical purposes there had been a death of societal organization
by the end of Phase C.
Decline of Population Size, the Death Phase D
Population increase abruptly cased on Day 560 after colonization. A few
mice born up until Day 600 survived past weaning. Between these times deaths
just slightly exceeded births. Beyond the time of the last surviving birth
on Day 600 the incidence of pregnancies declined very rapidly with no young
surviving. Last conception was about Day 920. With the increase in rate
of mortality accompanying senescence, the population has continued to decline
in numbers. By March 1 1972, the average age of survivors was 776 days,
over 200 days beyond menopause.
The last surviving male will
be dead on May 23, 1973,
1780 days after colonization.
On June 22 1972, there were only 122 (22 male, 100 female) survivors.
Projection of the prior few months of exponential decline in numbers indicates
that the last surviving male will be dead on May 23 1973, 1780 days after
colonization. The population will be, reproductively, definitely dead at
that time, although such death was predicted by 700 days after colonization.
This demise of a population contradicts prior knowledge which indicates
that when a population declines to a few remnant groups, some individuals
will reinitiate its growth.
Turning back to the end of phase C, the seeds for eventual destruction may
already be seen to have been sown. By midway in Phase C essentially all
young were prematurely rejected by their mothers. They started independent
life without having developed adequate affective bonds. Then as they moved
out into an already dense population many attempts to engage in social interaction
were mechanically disrupted by passage of other mice. Lastly, I have shown
(Calhoun 1963) that in proportion to the extent that the group size exceeds
the optimum, maximizing gratification from such interactions necessitates
a decrease in the intensity and duration of such behaviors. This fragments
otherwise more complex behaviors. As a result of these three processes
(failure to develop early social bonding, mechanical interference with developing
social behaviors and fragmentation of behaviors) maturation of the more
complex social behaviors such as those involved in courtship, maternality
and aggression failed. For females a clear example may be taken from a 2
cell universe studied in parallel with the 16 cell one detailed here. The
members of this population were killed 300 days after the inflection point
of the shift from Phase C to Phase D. Among these were 148 females born
within the last 50 days before the end of Phase C. At autopsy at a median
age of 334 days only I 8% had ever conceived (i.e. no placental scars in
the uteri of 82% of the females) and only 2% were pregnant (each of these
3 females had only one embryo as contrasted to the more normal S or more).
By this age most females in a normal population would have had five or more
litters, most of them successfully reared.
Male counterparts to these
non-reproducing females
we soon dubbed the
'beautiful ones'.
Male counterparts to these non-reproducing females we soon dubbed the
'beautiful ones'. They never engaged in sexual approaches toward females,
and they never engaged in fighting, and so they had no wound or scar tissue.
Thus their pelage remained in excellent ,condition. Their behavioural repertoire
became largely confined to eating, drinking, sleeping and grooming, none
of which carried any social implications beyond that represented by contiguity
of bodies.
Most of the last half of the population born in the 16 cell universe were
fully or largely like these non-reproducing females and these 'beautiful
ones' (males). As their formerly more competent predecessors gradually became
senescent, their already disrupted capacity for reproduction terminated.
At this time only the 'beautiful one' category of males, and their counterpart
females, remained at an age normally compatible with reproduction, but they
had long since failed to develop this capacity.
My colleague, Dr. Halsey Marsden (1972), conducted several studies during
the mid-third of Phase D in which he placed small groups of mice out of
these crowded populations into new universes at very low densities. All
groups exhibited nearly total loss of capacity for developing a structured
society or for engaging in the full repertoire of reproductive behaviors.
Even placing them with adequate sex partners of the opposite sex, that had
matured in uncrowded conditions, also gave very little indication of retention
of any adequate reproductive behavior.
Conclusion
The results obtained in this study should he obtained when customary causes
of mortality become markedly reduced in any species of mammal whose members
form social groups. Reduction of bodily death (i.e. 'the second death')
culminates in survival of an excessive number of individuals that have developed
the potentiality for occupying the social roles characteristic of the species.
Within a few generations all such roles in all physical space available
to the species are filled. At this time, the continuing high survival of
many individuals to sexual and behavioral maturity culminates in the presence
of many young adults capable of involvement in appropriate species-specific
activities. However, there are few opportunities for fulfilling these potentialities.
In seeking such fulfillment they compete for social role occupancy with the
older established members of the comununity. This competition is so severe
that it simultaneously leads to the nearly total breakdown of all normal
behavior by both the contestors and the established adults of both sexes.
Normal social organization (i.e. 'the establishment') breaks down, it 'dies'.
Young born during such social dissolution are rejected by their mothers
and other adult associates. This early failure of social bonding becomes
compounded by interruption of-action cycles due to the mechanical interference
resulting from the high contact rate among individuals living in a high
density population. High contact rate further fragments behavior as a result
of the stochastics of social interactions which demand that, in order to
maximize gratification from social interaction, intensity and duration of
social interaction must be reduced in proportion to the degree that the
group size exceeds the optimum. Autistic-like creatures, capable only of
the most simple behaviors compatible with physiological survival, emerge
out of this process. Their spirit has died ('the first death'). They are
no longer capable of executing the more complex behaviors compatible with
species survival. The species in such settings die.
For an animal so complex as man, there
is no logical reason why a comparable
sequence of events should not also
lead to species extinction.
For an animal so simple as a mouse, the most complex behaviors involve
the interrelated set of courtship, maternal care, territorial defence and
hierarchical intragroup and intergroup social organization. When behaviors
related to these functions fail to mature, there is no development of social
organization and no reproduction. As in the case of my study reported above,
all members of the population will age and eventually die. The species will
die out.
For an animal so complex as man, there is no logical reason why a comparable
sequence of events should not also lead to species extinction. If opportunities
for role fulfillment fall far short of the demand by those capable of filling
roles, and having expectations to do so, only violence and disruption of
social organization can follow. Individuals born under these circumstances
will be so out of touch with reality as to be incapable even of alienation.
Their most complex behaviors will become fragmented. Acquisition, creation
and utilization of ideas appropriate for life in a post-industrial cultural-conceptual-technological
society will have been blocked. Just as biological generativity in the mouse
involves this species most complex behaviurs, so does ideational generativity
for man. Loss of these respective complex behaviors means death of the
species.
Mortality, bodily death = the second death
Drastic reduction of mortality
= death of the second death
= death squared
=(death)2
(Death)2 leads to dissolution of social organization
=death of the establishment
Death of the establishment leads to spiritual death =loss of capacity to
engage in behaviors essential to species survival
=the first death
Therefore:
(Death)2=the first death.
Happy is the man who finds wisdom, and the man who gains understanding.
Wisdom is a tree of life to those who lay hold of her.
All her paths lead to peace. (Proverbs iii.13, 18 and 17, rearranged)
REFERENCES
Calhoun J B
(19,63) In: Physiological Mammalogy. Ed. W Mayer &
Ryan Gelder. Academic Press. New York; 1, I-I 117
(1967) In: Comparative Psychopathology. Ed. J Zubin.
Grune & Sirattoe,, New York; pp 1-51
(1969) Architectural Association Quarterly 1, No.3, 24-35
(1971) In: Behavior and Environment - The Use of Space:ace by
Animals and Men. Ed. A H Esser. Plenum, New York; pp 329-3117
Maruden H M (1972) In: Environment and the social Sciences:
Perspectives and Applications. Ed. J F Wohwill & D H Carson.
American Psychological Association, Washington, DC; pp 5-14
Marsden H M ,Calhoun J B & Ng LKY
(1971) Crowding and Maladaptation: Behavioral and
Neurochemical Observations. (Paper presented at the
Annual Meeting of the American Association for the
Advancement of Sciences, Philadelphia, December 1971
(unpublished)
Wigotsky Y W (1970) In: Engineering and the
Urban Crisis: Part 3. Urban Congestion,
Design News, Cahners, Denver, Col.; September 15, pp 48-60
|
|
|
| | | |
|
{kind=link}